|
Строение зрительного пути
Топографические особенности глазного дна
Топографические особенности глазного дна обусловлены значительными различиями в строении и функции сетчатки в зависимости от ее расположения, на основании которых на глазном дне выделяют центральную и периферическую зоны сетчатки и, кроме того, отдельные зоны в центральной сетчатке.
Если с помощью офтальмоскопа заглянуть через зрачок внутрь человеческого глаза, можно увидеть картину, изображенную на рисунке 6. Так выглядит центральная зона сетчатки здорового человека. На рисунке 7 схематично представлена топография и номенклатура центральной зоны сетчатки.
Термины фовеола, фовеа и макула появились вследствие особенностей гистологического строения сетчатки в каждой из этих зон. Фовеола - содержит только фоторецепторы, ганглиозные клетки в ней отсутствуют. Фовеа – зона, в которой сетчатка истончается до половины своей обычной толщины. Перифовеальная зона (макула или "желтое пятно") характеризуется высокой концентрацией колбочек и многослойным расположением ганглиозных клеток; наружная граница макулы определяется местом перехода к размещению ганглиозных клеток в один слой. Сам термин «задний полюс глаза» или «центральная сетчатка» появился в связи с необходимостью локализации патологических изменений на глазном дне при заболеваниях глаз. А в общем он является синонимом термина «макула».
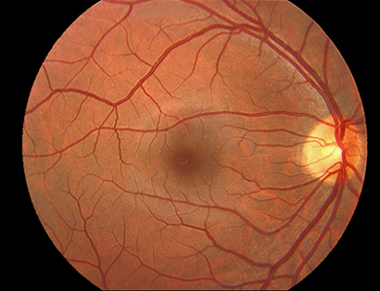
Рис. 6. Нормальное глазное дно правого глаза человека

Рис. 8. Световая микроскопия макулы
|
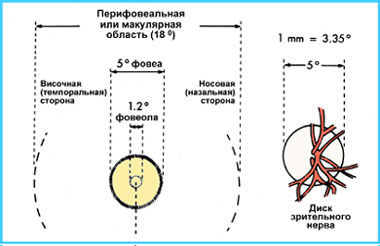
Рис. 7. Топография центральной зоны глазного дна
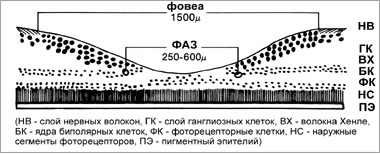
Рис. 9. Фовеа, фовеальная аваскулярная зона (ФАЗ) и фовеола
|
1.2. Сенсорная часть сетчатки
Сенсорная часть сетчатки располагается за слоем пигментного эпителия и выстилает глазное яблоко изнутри. Она представляюет собой тонкую прозрачную оболочку, содержащую чувствительные к свету клетки, которые и превращают световую энергию в нервные импульсы.
На поперечном гистологическом срезе сенсорная часть сетчатки выглядит многослойной.
|
Слои сенсорной части сетчатки
|
|
1. Пигментный эпителий
2. Слой фоторецепторов (палочек и колбочек)
3. Наружная пограничная мембрана
4. Наружный ядерный слой
5. Наружный плексиформный (сетчатый) слой
|
6. Внутренний ядерный слой
7. Внутренний плексиформный (сетчатый) слой
8. Слой ганглиозных клеток
9. Слой нервных волокон
10. Внутренняя пограничная мембрана
|
Гистологическое разделение сетчатки на слои, однако, не полностью отражает ее функциональной направленности - восприятие и переработку света. Это обусловлено тем, что в состав одиного гистологического слой могут входить клетки разных функцинальных типов, и наоборот - некоторые клетки могут располагаться на протяжении нескольких гистологических слоев. Поэтому с функциональной точки зрения сетчатку целесообразно подразделить на нейронные системы, выполняющие ту или иную функцию в процессе восприятия, переработки зрительной информации и ее передачи по зрительному пути.
В сетчатке выделяют две нейронные системы - вертикальную и горизонтальную, состоящие из 6 типов нейронов.
Вертикальная представлена иерархией трех клеточных элементов: 1) фоторецепторные клетки (первый нейрон зрительного пути), 2) биполярные клетки (второй нейрон) и 3) ганглиозные клетки. Именно в этой цепи нервный импульс, сформированный в фоторецепторе под воздействием света, передается головному мозгу посредством зрительного нерва.
Горизонтальная система нейронных связей состоит из нейронов, определяющих связь между фоторецепторами, биполярными и ганглиозными клетками в плоскости сетчатки. К этим нейронам относятся горизонтальные (в наружном плексиформном слое), амакриновые (во внутреннем плексиформном слое) и межплексиформные клетки.
Кроме вышеуказанных типов нервных клеток, в состав сенсорной сетчатки входят также глиальные элементы - клетки Мюллера, фиброзные и протоплазматические астроциты, микроглия и олигодендроциты.
Ниже будут детально представлены все клеточные типы, входящие в состав сенсорной сетчатки, а также рассмотрена функциональная анатомия зрительного пути.

Рис. 10. Схематическое изображение разных видов нейронов сетчатки
(ибк - инвагинированные биполяры колбочек, пбк - плоские биполяры колбочек, бск - биполяры "синих" колбочек, пдбк - плоские диффузные биполяры колбочек, бк - биполяры палочек, гк - горизонтальные клетки, ха - холинэргические амакриновые клетки, ипк - интерплексиформные клетки, ак - амакриновые клетки, идбк - инвагинированные диффузные биполяры колбочек)
1.2.1. Фоторецепторы (колбочки и палочки)
Сетчатка содержит два вида фоторецепторных клеток - палочки и колбочки, которые различаются функционально и анатомически. Колбочки отвечают за высокое пространственное разрешение (остроту зрения) и цветовое зрение при высокой освещенности (фотопическое зрение), тогда как палочки обеспечивают высокую монохроматическую чувствительность глаза (хоть и с относительно низким пространственным разрешением) при низкой освещенности (скотопическое зрение).
Анатомически слой колбочек и палочек является самым наружным слоем сенсорной сетчатки. Он состоит из цитоплазматических выростов фоторецепторных клеток, которые представляют собой высокоспециализированные светочувствительные нейроэпителиальные клетки. Тела фоторорецепторов располагаются в плоскости наружной пограничной мембраны, а их апикальные отростки (внутренние членики или сегменты) лежат снаружи этой мембраны.
Фоторецепторы в сетчатке плотно прилежат друг к другу и пространственно ориентированы вдоль зрительной оси, что обеспечивает детальный анализ поля зрения. Любое изменение расположения фоторецепторов приводит к нарушению зрения - нарушение ориентации вдоль зрительной оси приводит к искажению зрительного образа (метаморфопсии), при появлении пространств между фоторецепторами развиваются микропсии (уменьшение изображения предметов).
В сетчатке насчитывают от 77,9 до 107,3 млн. (в среднем 92 млн.) палочек и 4,1-5,3 млн.(в среднем 4,6 млн.) колбочек.
Фоторецепторы распределяюся закономерным образом в виде мозаики. В области желтого пятна лежат только колбочки. Вне желтого пятна колбочки кольцевидно окружены палочками.
Существуют индивидульные различия плотности колбочек и палочек в зависимости от топографического отдела сетчатки. Наибольшее разнообразие плотности выявлено вблизи центральной ямки и у зубчатой линии, наименьше - в средней части сетчатки и по периферии.
Плотность колбочек максимальна в области центральной ямки (199 000 колбочек в мм ² ). По мере удаления от центральной ямки плотность колбочек существенно уменьшается. Степень этого уменьшения различна в зависисмости от направления - с назальной стороны плотность колбочек на 40-45% выше, чем с темпоральной. В периферических отделах сетчатки плотность колбочек опять возрастает. Считается, что пространственное расположение колбочек в области желтого пятна определяет разрешающую способность глаза .
Наименьшее расстояние между колбочками выявлено в центральной ямке. Этим обусловлена наибольшая разрешающая способность сетчатки именно в этой области. Однако, есть сведения, что данные психофизиологических исследований остроты зрения не полностью соответствуют анатомическим данным. Вероятно, в формировании остроты зрения большое значение имеют другие факторы. Единственная зона в сетчатке, где функциональная острота зрения совпадает с анатомической расположена между 0,2 и 2,0 град.
Человек рождается с неполностью дифференцированным желтым пятном. Поэтому у новорожденных острота зрения на два порядка ниже, чем у взрослых. В ближайшее время после рождения колбочки, палочки и клетки пигментного эпителия перемещаются к центру желтого пятна. Плотность колбочек у человека нарастает после рождения вплоть до 5-8 летнего возраста. Однако наиболее важным фактором, определящим низкую остроту зрения у новорожденных, является не плотность колбочек в центральной сетчатке, а неполная дифференциация желтого пятна.

Рис. 11. Плотность палочек и колбочек вдоль горизонтального меридиана
|
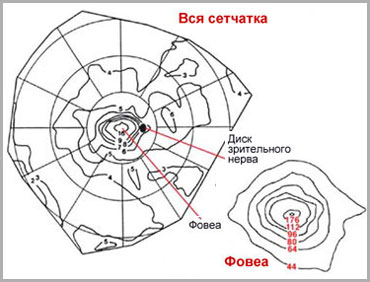
Рис. 12. Топографическая карта плотности колбочек в сетчатке человека (по Curcio et al., 1987)
|
Плотность палочек. В центральной области желтого пятна диаметром 0,35 мм (1,25 град) палочки отсутствуют. Область самой высокой концентрации палочек в сетчатке имеет форму горизонтального эллипса, который несколько расширяется в носовом направлении кверху. Далее по направлению к периферии плотность палочек медленно снижается. С назальной стороны плотность палочек на 20-25% выше, чем с височной стороны.
Строение фоторецепторов. Внутренние и наружные сегменты фоторецепторов являются местом трансформации световой энергии в нервный импульс. Оба вида фоторецепторов - палочки и колбочки - имеют сходную организацию, которая различается в определенных деталях. В целом в каждой фоторецепторной клетке можно выделить следующие части (по направлению от хориоидальной стороны кнутри к стекловидном телу) (Рис. 11):
1) наружный и внутренний сегменты, которые вместе формируют апикальный отросток фоторецептора; отросток колбочки широкий, но заостренный на конце (в соответствии с названием), тогда как отросток палочки имеет строго цилиндрическую форму. Наружный и внутренний сегменты связаны между собой тонким перешейком.. Внутренний сегмент подразделяется на наружную часть - эллипсоид и внутреннюю - миоид.
2) наружные волокна (их вид зависит от типа фоторецепторов и области их локализации в сетчатке) - короткий отросток, соединяющий наружный и внутренний сегменты фоторецепторов с телом клетки;
3) тело клетки (или сома), содержащее ядро;
4) внутренние волокна, соединяющие тело клетки с синаптическими окончаниями фоторецептора -
5) синаптическое окончание - "ножка" палочки и сферула колбочки ; здесь находится зона синаптического контакта фоторецептора с прилежащими биполярными и горизонтальными клетками

Рис. 13. Схематическое изображение строения палочки (А) и колбочки (В)
Наружные сегменты палочек и колбочек. Ультраструктурно колбочки и палочки очень похожи. Их наружные сегменты выглядят как четко определяемые регулярные серии дисковидных уплощенных мешочков, упакованных в фоторецепторе подобно монетным столбикам и окруженных клеточной мембраной. Наружный конец сегментов прилежит к клеткам пигментного эпителия. Диски представляют собой плоские пузырьки (везикулы), отделенные друг от друга междисковым пространством низкой электронной плотности. Количество дисков в наружном сегменте палочек составляет 600-1000, в колбочках их число выше - 1000-1200. Никаких специализированнх контактов между дисками, а также дисками и цитоплазматической мембраной в палочках не обнаружено. Напротив, диски колбочек соединены между собой и прикрепляются к цитоплазматической мембране.
Наружный членик обоих видов фоторецепторов постоянно обновляется за счет постоянного образования дисков на его конце, обращенном к телу клетки, и безостановочного их продвижения по направлению к пигментному эпителию, где происходит фагоцитоз "старых" дисков клетками пигментного эпителия. Взамен фагоцитируемым дискам постоянно образуются новые. Таким образом, процесс образования, старения и утилизации дисков является непрерывным, по сути образуя цикл.
С обновлением наружного членика фоторецептров постоянно регенерируют расположенные на дисках зрительные пигменты - специфические вещества, запускающие фотохимическую реакцию, в результате которой происходит преобразование световой энергии в электрические импульсы. В дальнейшем, нервные импульсы обрабатываются на уровне сетчатки и передаются по зрительному нерву в кору головного мозга.

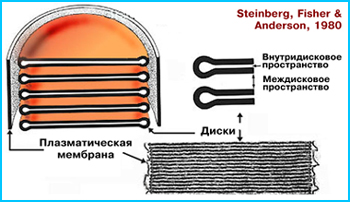
Рис. 14. Схема образования и продвижения дисков в наружных сегментах фоторецепторов
|

Рис. 15. Наружный сегмент фоторецептора и его дисковая структура (А) и схематическое представление родопсина в дисках (В)
(по Carlos Rozas, Chiliе)
|
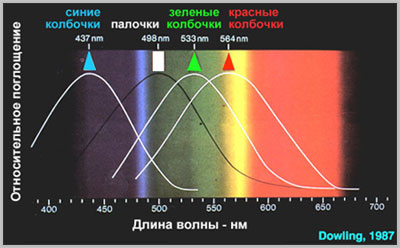
Восприятие света в фоторецепторах и преобразование его в нервный импульс начинаются с активации фотохимической стереоизомеризации зрительного пигмента, расположенного на дисках наружного сегмента фоторецептора. В сетчатке человека выявлено четыре типа зрительных пигментов. Один тип обнаружен в палочках (родопсин) и три в колбочках (йодопсин). В зависимости от спектральных особенностей поглощения световой энергии колбочковые пигменты разделяются на чувствительные к красной (570 нм), зеленой (540 нм) и синей (440 нм) частям спектра. Соответственно, колбочки содержащие каждый из этих пигментов, обозначаются как L- (длинноволновые 570 нм), M- (средневолновые - 540 нм) и S- (коротковолновые - 440 нм) колбочки.
В настоящее время морфологическими методами дифференциируют средне- и длинноволновые колбочки у некоторых рептилий, земноводных, рыб и птиц. У приматов и человека возможна дифференциация только коротковолновых "синих" колбочек (S-колбочки) от остальных. С этой целью используются специальные антитела, которые позволяют избирательно выделить содержащийся в колбочках зрительный пигмент, чувствительный к коротковолновой части спекта ("синий" пигмент).
У "синей" колбочки наружный членик более длинный и большего диаметра. Кроме того, "синие" колбочки не столь равномерно и закономерно распределены в сетчатке.
Рис. 16. Пики спектральной чувствительности трех видов колбочек и палочек в сетчатке приматов
(Brown & Wald, 1963)
В области желтого пятна "синие" колбочки встречаются редко (3-5% от общего числа фоторецепторов) и лежат изолированно либо полностью отсутствуют в зоне пика наибольшей плотности колбочек. Диаметр этой зоны равен 1000 мкм (0,35 градуса). Пик плотности "синих" колбочек (более 2000 в мм) выявлен в области фовеального склона (около 12%) в зоне шириной 0,1-0,3 мм.
Внутренние сегменты фоторецепторов.
Ультраструктурная организация внутренних сегментов палочек и колбочек одинаковая, за исключением того, что в колбочках значительно больше митохондрий. Внутренние сегменты обоих видов фоторецепторов имеют цилиндрическую форму и состоят из двух частей - наружной (эллипсоид) и внутренней (миоид). В эллипсоидной части содержится большое количетво митохондрий, гладкая эндоплазматическая сеть, свободные рибосомы, нейротрубочки и гранулы гликогена. Миоидная область является центром белкового синтеза. По этой причине в ней содержится шероховатая эндоплазматическая сеть, аппарат Гольджи, микротрубочки, микрофиламенты и гранулы гликогена. Таким образом, основной функцией этой части фоторецептора является обеспечение метаболических и синтетических функций клетки.
Наружная поверхность миоидной части фоторецепторов покрыта воловоподобными цитоплазматическими отростками клеток Мюллера, формирующих "корзины Шульца". Благодаря этому никакого контакта между клетками нет. Отростки клеток Мюллера участвуют также в регуляции состава внеклеточной среды фоторецепторов и служат для жесткой пространственной фиксации палочек и колбочек.
Наружные волокна - дендритоподобные отростки, соединяющие внутренние сегменты с телом фоторецептора. Их длина различается в палочках и колбочках в зависимости от расположения в сетчатке. Содержат множество микротрубочек и несколько митохондрий.
Тело клетки (сома) - эта область включает в себя ядро, размер которого в палочках меньше. Вокруг ядра цитоплазма формирует относительно узкую щель, включающую небольшое число органелл.
Внутренние волокна - аксоноподобные образования, содержащие митохондрии, везикулы, свободные рибосомы, нейрофиламенты и микротрубочки. В области желтого пятна и вокруг него внутренние волокна колбочек расположены косо, а за пределами макулы образуют почти горизонтальный слой (волокна Хенле).
Cферулы палочек и "ножки" колбочек по сути являются местом синаптических контактов фоторецепторов с биполярными и горизонтальными клетками, а также друг с другом.
Внутренние волокна (аксоны) палочек заканчиваются в наружном сетчатом (плексиформном) слое грушевидным или овальными окончаниями - сферулами. Аксоны колбочек образуют более широкие, уплощенные "ножки". Оба типа окончаний формируют множественные соединения с биполярными и горизонтальными нейронами, нейриты которых приходят из внутреннего ядерного слоя и, во многих случаях, глубоко погружаются в основание сферулы или "ножки".
Cферулы палочек . Синаптический комплекс палочек состоит из самой сферулы, синаптической ленты и постсинаптических отростков, принадлежащих горизонтальным или биполярным клеткам. Кроме того, сферулы посредством щелевых соединений соединяются со сферулами или "ножками" других палочек и колбочек. Основание сферулы глубоко вогнуто и вмещает в своей полости триаду - характерное синаптическое устройство, содержащее многочисленные синаптические пузырьки и синаптическую ленту, направленную на комбинацию из одного или больше дендритов центральных биполяров, окруженных двумя аксонами HI горизонтальных клеток. Это устройство обеспечивает комплексное синаптическое взаимодействие между тремя видами клеток за счет возбуждающих и тормозящих воздействий. Синаптическая летна представляет собой перпендикуляр, проходящий через пресинаптическую мембрану и состоящий из трех электронноплотных слоев толщиной 12 мкм каждый. Сферулы палочек содержат только две синаптические ленты, которые ассоциируются с двумя боковыми элементами - окончаниями аксонов горизонтальных клеток и двумя дендритами биполярных клеток палочек.
С одной сферулой палочки может входить в контакт несколько различных горизонтальных клеток (1-4) и 4 биполярные клетки. В то же время каждая биполярная клетка контактирует с 50 палочками вне фовеолы и несколькими сотнями палочек - по периферии. Эти различия в характере межнейронных связей соответствуют различиям в разрешающей способности зрительной системы.

"Ножки" колбочек. Это более обширный комплекс: у приматов он объединяет в себе 20-30 синаптических вдавлений на "ножку". Каждое синаптическое вдавление объединяет в себе три нейрона - типичная триада, состоящая из аксона биполяра колбочки ("ON"-биполяры) и двух обнимающих его дендрита разных горизонтальных клеток. Один аксон может входить в контакт с одной и то же колбочкой в 10-25 различных точках. Одна "ножка" колбочки контактирует с одной биполярной клеткой, которая, в свою очередь, имеет контакты с 6-8 горизонтальными клетками. Такая ножка имеет также множество небольших поверхностных вдавлений (так называемых базальных соединений), которые контактируют с плоской диффузной колбочковой биполярной клеткой ("OFF"-биполяры). Подобный тип синапсов биполярных клеток формируется сразу с 6 колбочками. Каждая "ножка" может иметь соединения с двумя карликовыми и 15 диффузными биполярными клетками.
Рис. 17. Схематическое изображение синаптических тел колбочек и палочек
Кроме биполярных и горизонтальных клеток, фоторецепторы контактируют и между собой. Происходит это благодаря щелевидным контактам. От "ножки" колбочки отходят тонкие отростки (телодендриты), которые подходят к сферулам палочек и "ножкам" других колбочек и формируют так называемый "электрический контакт", где передача импульса осуществляется без нейротрансмиттера. На одной сферуле палочки определяется 3-5 подобных контактов. Одна "ножка" колбочки может иметь до 10 контактов с соседними палочками. "Ножки" S-колбочек не содержат такого большого количества контактов. По этой причине "синие" колбочки довольно изолированы.
Функциональное значение приямой электрической связи между различными типами фоторецепторов не совсем понятно. Первоначально предполагалось, что такие связи разрушают пространственную интеграцию фоторецепторов и, соответственно, возможность анализа работы цветового зрения, смешивая информацию, полученную от палочек и колбочек. Тем не менее, на основании многих исследований установлено, что колбочки, благодаря этим связям, могут нести информацию палочек.
|
